Neurons are talkers. They talk to muscles and tell them to move bones. They speak to the heart and ask it to speed up or slow down. When chased by a pterodactyl, the burst of energy came from a neuron yelling at the adrenal glands to bathe the body in adrenalin.
However, neurons also know when to listen. They listen to the eyes, skin, and nose. Furthermore, the heartburn one feels after eating 27 chili cheese dogs comes from listening to neurons.
So, how do neurons speak and listen to other cells? Neurons talk to each other at the synapse.
Presynaptic and Postsynaptic Neurons
A synapse is a location where a neuron interacts with another cell-e.g., the dendrite of one neuron and the terminal endings of another neuron. A postsynaptic neuron is a neuron that receives a stimulus. Postsynaptic dendrites receive stimuli from the environment, another cell, or presynaptic neurons. Presynaptic neurons stimulate postsynaptic neurons by
- releasing chemical messengers (neurotransmitters) into the gap between the two cells.
- ion exchange.

Types of Synapses
Electrical Synapses
Gap junctions connect electrical synapses. A gap junction is a union of channel proteins between two neurons that speeds up communication. Ions from a presynaptic neuron flow through gap junctions into the postsynaptic neuron. The influx of ions into the postsynaptic neuron will cause a graded potential.
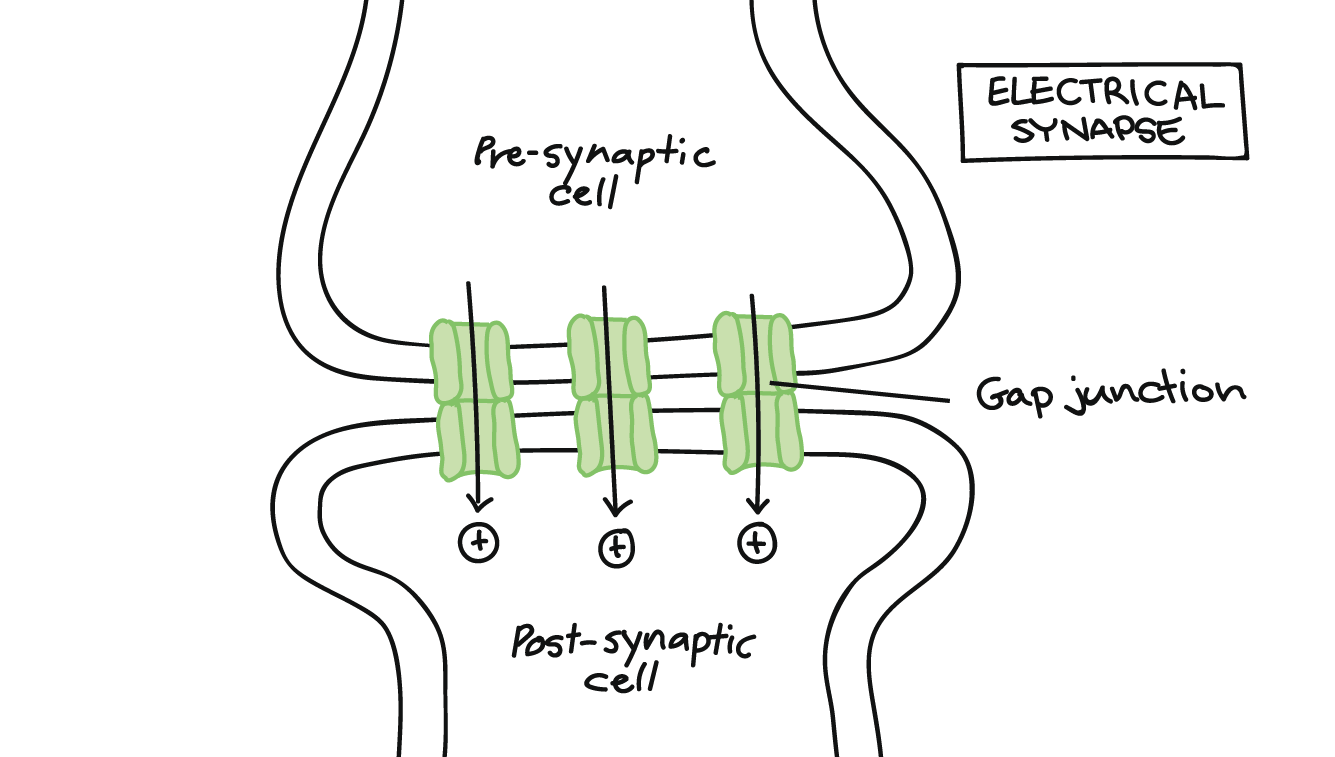
Electrical synapses transmit information faster and misfire less than chemical synapses. Their location is in areas of the nervous system where neuron synchrony is paramount. Neuron synchronization occurs when groups of neurons fire action potentials simultaneously and are an integral part of neural communication. For example, a network of electrical synapses in the thalamus filter and synchronize stimuli. Without synchronization of the synapses, seizures may develop.
Chemical synapses
Unlike electrical synapses, chemical synapses do not form gap junctions. The synaptic cleft is a small gap between the presynaptic and postsynaptic neurons. The presynaptic neuron releases chemical messengers called neurotransmitters into the synaptic cleft. The neurotransmitters diffuse across the synaptic cleft and bind to their target receptor protein (protein with the matching shape). The binding of the neurotransmitter will lead to an inhibitory or excitatory graded potential.
Chemical synapses are greater in number, slower, and more error-prone than electrical synapses. However, chemical synapses are more versatile and adaptable compared to electrical synapses. Chemical synapses can be regulated by
- the number of neurotransmitters released from the presynaptic neuron.
- how receptive the postsynaptic membrane is to the neurotransmitters.
- the amount of neurotransmitter receptor proteins on the postsynaptic cell’s membrane.
- increasing or decreasing the number of synapses between neurons.
The constant evolution of chemical synapses is how we learn and respond to our environment, contributing to our primary biological purpose: survival and reproduction.

Neural Plasticity
The brain is a very dynamic organ and is in constant flux. Every millisecond, new synapses form, and old synapses disconnect, a process called neural plasticity. Neural plasticity happens throughout life but is most active in the first 20-30 years and comprises two stages; synaptic formation and synaptic pruning.
Synaptic formation is most active in the first two years of life. By the age of two, synaptic connections reach their apex. However, there are too many synapses, so the brain begins synaptic pruning by removing the inactive and strengthening the active synapses (click here and here for more information on synaptic pruning). Synaptic or neural pruning primarily occurs during REM sleep (the dream phase of the sleep cycle). This may be why most dreams appear to be a potpourri of random events because they may be nothing more than fragments of synaptic plasticity. Synaptic pruning is one of the many reasons why 8-9 hours of sleep a day is paramount.



By 30, a person will have half as many synapses as they did when they were two years old. However, a 30-year-old brain’s synapses are stronger and more efficient than a two-year-old brain’s. For example, assume a person is trying to solve a puzzle with 30 people. The 30 people are simultaneously giving their advice on how to solve the puzzle. However, each person’s input is vague, and it is difficult to discern which information to use because they speak simultaneously. This chaotic situation will make it difficult for the person to find a solution to the puzzle. Now, imagine the person solving the same puzzle with two knowledgeable individuals. Listening to two well-informed people will allow the person to solve the puzzle faster. Why? Working with two knowledgeable people is easier than working with 30 less knowledgeable people. The same principle is true for neurons. Fewer inefficient neurons will allow the brain to function better.

Why We Dream
The significance of dreams and their meaning has been a topic of discussion that precedes written history. The hypotheses on dreams have a broad range, from our subconscious predictions to random neuronal noise. However, recent research has focused on two interesting hypotheses.
First, we do much neural pruning during REM sleep (dream sleep), and dreams appear to strengthen used neural connections and prune weak connections.
Second, a new study has developed a fascinating hypothesis that suggests why most vertebrates dream. Neural placidity is a rapid process that happens within minutes to hours. When we sleep, our eyes are closed, and we do not receive stimuli for eight hours. During sleep, the lack of stimuli will break neural connections in our brain’s visual centers and form new connections in the other sensory areas. For example, blind individuals can read braille because their brains are wired to use touch to read. If you were to put your finger over the braille, you probably could not decipher the different letters. However, if you were to blindfold yourself for eight or more hours, your ability to distinguish braille letters will improve. This happens because your brain no longer receives visual sensory information and now uses tactile information to read. Therefore, neural pruning occurs, weakening the visual connections and strengthening the tactile sensations. To prevent this from happening while we sleep, we dream 2-3 minutes every 90 minutes so our visual brain centers’ important connections remain intact over our slumber.
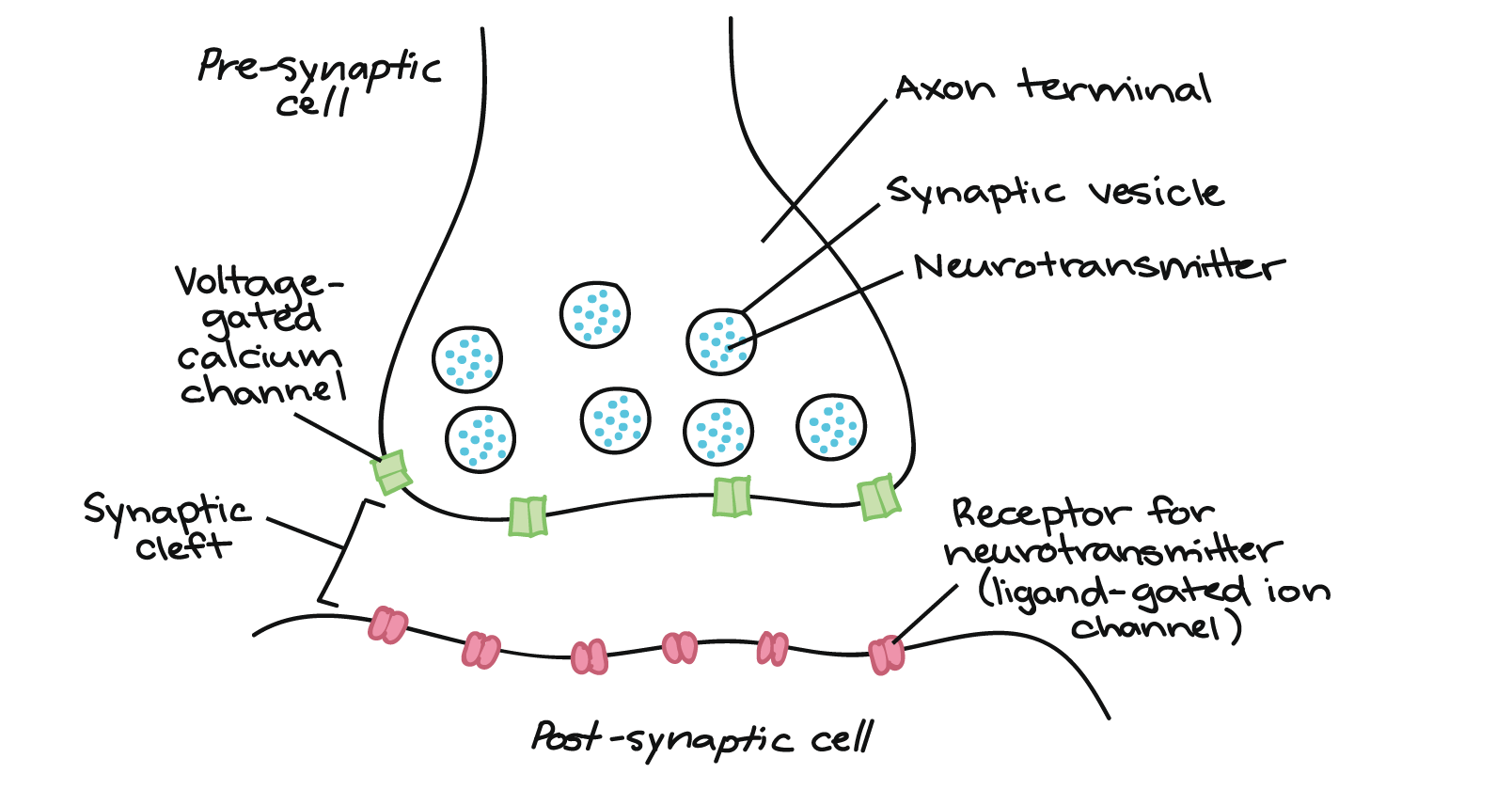
Transmission Across a Chemical Synapse
The terminal endings of a neuron contain vesicles filled with neurotransmitters. A vesicle is a membrane-bound sack that transports neurotransmitters or peptide hormones within a cell. The terminal ends are rich in mitochondria because a neuron uses exocytosis (active transport) to release neurotransmitters.
Each neuron makes and releases one type of neurotransmitter. There are many different types of neurotransmitters, but they all inhibit or excite the postsynaptic cell. For example, the neurotransmitter glutamate excites postsynaptic cells, and the neurotransmitter GABA inhibits postsynaptic cells. However, most neurotransmitters can excite or inhibit postsynaptic cells.
Synaptic transmission occurs as follows:
- When an action potential reaches the terminal endings of a presynaptic neuron, it stimulates calcium channel proteins embedded in the terminal endings’ membranes to open, and Ca2+ diffuses into the cell
- The influx of Ca2+ depolarizes the terminal ends and causes the synaptic vesicles to release their neurotransmitters via exocytosis into the synaptic cleft
- The neurotransmitters diffuse across the synapse and attach to their target receptor protein on the postsynaptic cell’s dendrite
- The attachment of the neurotransmitter will depolarize (excite) or hyperpolarize (inhibit) the postsynaptic cell’s dendritic membrane
- Enzymes in the synaptic cleft destroy the neurotransmitters, or the neurotransmitters return to the presynaptic neuron

EPSPs and IPSPs
There are two types of graded potentials: excitatory postsynaptic potentials (EPSPs) and inhibitory postsynaptic potentials (IPSPs). EPSPs occur when neurotransmitters stimulate the opening of sodium (Na+) channel proteins on the postsynaptic neuron’s dendritic membrane. The influx of Na+ depolarizes the postsynaptic membrane, and an EPSP will travel down its dendrite towards the axon hillock. IPSPs happen when neurotransmitters the opening chloride (Cl–) or K+ channel proteins on the postsynaptic cell membrane. The influx of Cl– or the efflux of K+ will hyperpolarize the postsynaptic neuron, and an IPSP will move toward the axon hillock.


Summation of Graded Potentials
Graded potentials lose strength as they travel down dendrites. So, it is rare that a single graded potential will have the strength to meet the threshold potential. Instead, a neuron either goes through spatial summation or temporal summation. Spatial summation occurs when a neuron receives EPSP and IPSP stimuli from two or more neurons. Temporal summation occurs when a postsynaptic neuron gets a rapid succession of EPSPs from a single neuron. If the spatial or temporal summation of EPSPs and IPSPs at the axon hillock reaches the threshold potential, the postsynaptic neuron will fire an action potential.



A simple equation can quantify the summation of graded potentials.
(Number of EPSPs x strength of EPSPs in mV) + (number of IPSPs x strength of IPSPs in mv) = summation of graded potentials at the axon hillock.
Let us use the formula above to solve the following:
Neuron A has a resting membrane potential of -70 mV and a threshold potential of -55 mV. The neuron receives stimulation from 32 presynaptic neurons. The axon hillock receives the following graded potentials and their voltage: 22 ESPSs of 2 mV each and 10 IPSPs of -3 mV. What is the summation of the graded potentials? Will Neuron A receive the stimulus it needs to reach its threshold potential?
(22 EPSPs x 2 mV) + (10 IPSPs x -3 mV) =
44 mV + -30 mV = 14 mV
14 mV is the summation of graded potentials, which is not enough stimulus to reach the threshold potential of -55 mV.
The axon hillock similarly sums up stimuli. However, a neuron’s axon hillock usually receives many, sometimes thousands, of EPSPs and IPSPs per millisecond and sums them in that time frame. (This is faster than you can put pen to paper or finger to keyboard.)